
PBJ | 华农夏瑞课题组发现荔枝miR482/2118靶向非编码PHAS基因产生phasiRNA调控GID1参与GA信号传导
时间:2023-11-17 阅读量:313
2023年11月15日,华南农业大学夏瑞课题组在Plant Biotechnology Journal上在线发表了题为“MicroRNA482/2118 is lineage-specifically involved in gibberellin signaling via the regulation of GID1 expression by targeting noncoding PHAS genes and subsequently instigated phasiRNAs”的研究论文,该研究通过对荔枝中microRNA482/2118(miR482/2118)触发的PHAS基因分析发现了一条物种特异的调控通路——荔枝miR482/2118通过靶向非编码的PHAS基因产生phasiRNA调控GID1的表达参与赤霉素信号传导。

miR482/2118广泛存在于种子植物中,是一个古老且相对保守的22-nt miRNA超家族,可以通过触发靶基因产生大量的phasiRNA。这些phasiRNA一方面可以顺式作用于其前体基因加强miR482/2118的基因沉默作用,另外一方面可以通过反式作用靶向其它基因,从而扩大miR482/2118沉默作用的效应。在双子叶植物中,miR482/2118主要通过靶向NBS-LRRs家族产生phasiRNA调控植物的抗病过程。但是,随着研究深入,在双子叶植物中发现miR482/2118也进化出了一些物种或谱系特异的新功能或者亚功能,这说明miR482/2118的功能在一些双子叶植物中得到了扩张。
荔枝,无患子科常绿果树,是中国南方重要经济果树。结合新发表的高质量基因组,本研究通过对荔枝miR482/2118成员及其靶标进行综合鉴定后深入挖掘和解析其中的一条涉及到赤霉素信号传导的新功能途径。结合多组学测序、基因工程和生物分子学实验最终揭示了荔枝miR482/2118在该通路通过级联触发phasiRNA介导赤霉素信号传导的受体GID1的表达参与赤霉素信号传导的调控网络。研究结果简述如下:
01
荔枝miR482/2118家族及其靶基因的重新鉴定
结合新发布的高质量的荔枝基因组,本研究在荔枝中共鉴定了13个LcMIR482/2118成员,其中有两个成员产生的miR482/2118的成熟序列完全相同。因此,通过对12个不同的Lc-miR482/2118的成熟序列进行比对,发现分别有六个成员属于miR482和miR2118亚型,其中miR482亚型在第一个位置比对上而miR2118亚型有两个碱基的位移(图1a)。相比于星链的序列,Lc-miR482/2118成熟链更加保守,且星链和成熟链形成不对称的错配使得第10个碱基的位置形成了一个气泡结构,导致Lc-miR482/2118成熟链长度为22-nt(图1b)。这些Lc-miR482/2118成员分布在六条染色体上,其中在6,7,14号体上成簇出现(图1c)。本研究中共鉴定了23个Lc-miR482/2118的PHAS靶基因(图1d),这23个PHAS靶基因的分布也显示出染色体上的聚集现象,例如,在染色体4上存在一个由七个靶基因组成的超级簇,其中所有基因都是与抗病性和抗逆性相关的基因(图1c)。其中除主要靶向NBS-LRR家族参与抗病的PHAS基因外,Lc-482/2118还触发一些注释为激酶、激素应答蛋白、钙离子传输蛋白等的PHAS基因产生phasiRNA(图1e)。
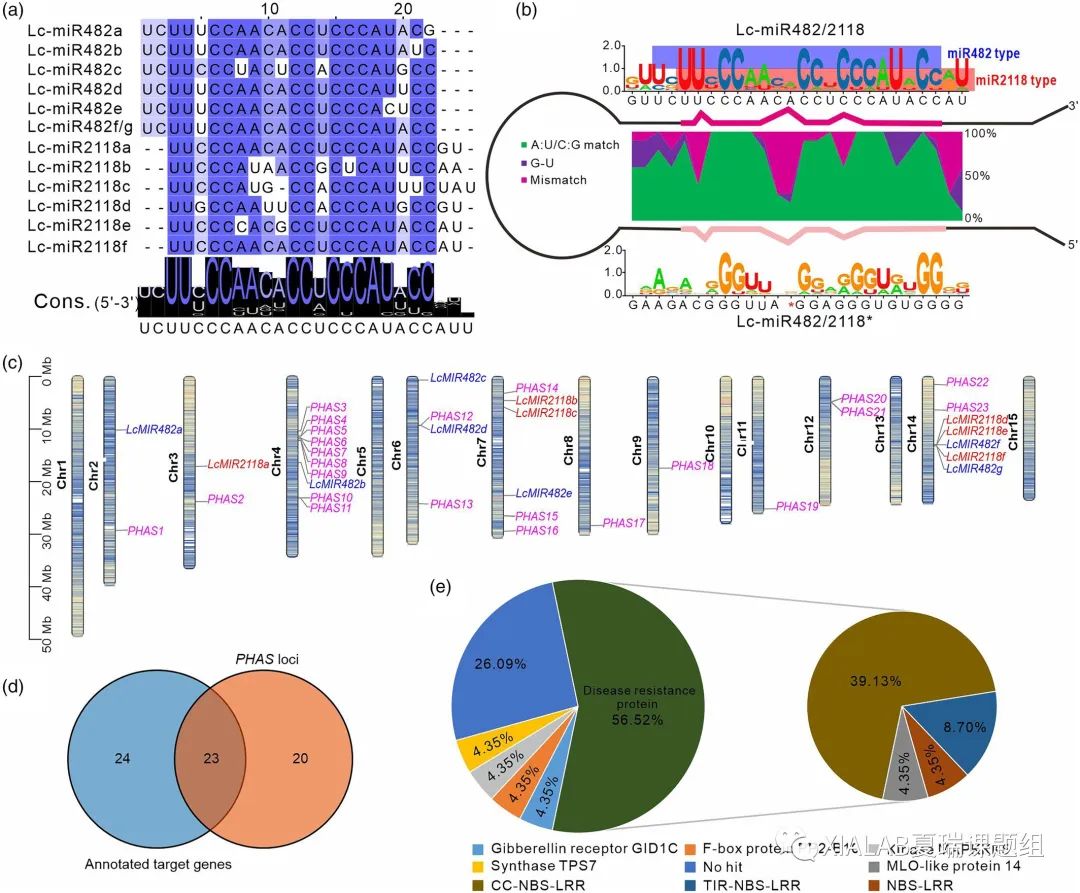
图1 荔枝miR482/2118家族及其靶基因的重新鉴定
02
Lc-miR482/2118通过触发非编码PHAS基因产生phasiRNA抑制赤霉素信号传导受体LcGID1实现新功能化
通过对Lc-miR482/2118的PHAS靶基因的分析,本研究发现荔枝miR482/2118可能进化出一些谱系和物种特异性功能(图1e)。对荔枝靶向的PHAS基因进行进一步的分析时,PHAS19基因(LITCHI007943)引起了我们的注意。PHAS19位于11号染色体上,具有5个外显子,通过降解组数据和5'RLM-RACE证明Lc-miR482/2118靶向切割LITCHI007943基因随后产生大量的phasiRNA(图2a和2b)。LITCHI007943全长转录本在核苷酸水平上显示出与赤霉素(gibberellin,GA)受体基因LcGID1具有高度序列相似性(图2c),但是通过编码性分析发现LITCHI007943是一个长链非编码RNA基因而非GID1同源基因,其预测最长的ORF与GID1结构域具有高度同源性(图2d),与LITCHI007943以及上产生phasiRNA的区域重叠(图2a和2c)。
我们发现,这些由LITCHI007943产生的phasiRNA能够在荔枝中反式切割LcGID1同源物(图2e和2f),这种作用类似于拟南芥中的反式作用基因(TAS)——一类非编码且产生tasiRNA反式作用下游基因的基因,因此我们将LITCHI07943命名为荔枝中的TAS类似基因1(LcTASL1)。
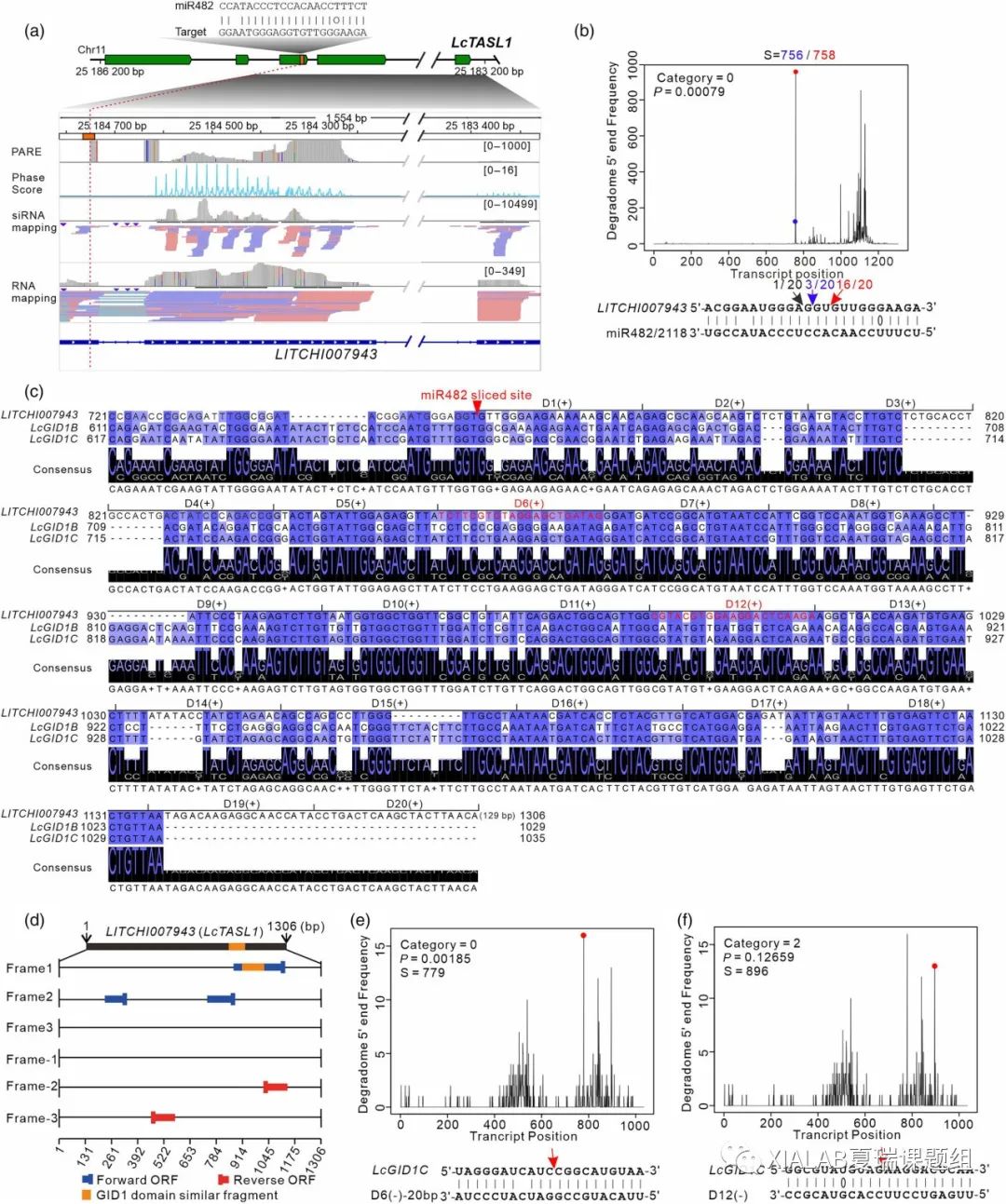
图2 Lc-miR482/2118s靶向一个长链非编码RNA基因LcTASL1触发phasiRNA产生调控LcGID1
03
LcTASL2产生级联phasiRNA增强miR482/2118-LcTASL1对LcGID1的作用
在已鉴定的PHAS基因位点中,我们注意到有一个与LcTASL1相邻的基因LITCHI007944(图3a)。序列分析显示,该基因是一个与LcGID1和LcTASL1具有很高序列相似性的长链非编码RNA基因(图3b)。与LcTASL1不同的是,在LITCHI007944上不存在miR482/2118的靶位点(图3a),这表明LITCHI07944没有被miR482/21118直接靶向。其phasiRNA的产生可能是由LcTASL1产生的phasiRNA触发的,因为它们的高序列相似性(50.03%)(图3b)。与LcTASL1类似的是,由LITCHI07944产生的phasiRNA也以反式靶向LcGID1(图S9);因此将LITCHI007944命名为LcTASL2。此外,两个LcGID1基因也产生了大量的phasiRNA(图3c和3d)。
总之,我们发现,在荔枝中,miR482/2118进化出一条新的通路——通过靶向LcTASL1产生phasiRNA调节参与GA信号传导通路的LcGID1,同时,LcTASL1产生的phasiRNA靶向的LcTASL2产生phasiRNA以增强对LcGID1基因的沉默作用(图3e)。此外,我们发现miR482/2118-TASL-GID1通路中TASL1和TASL2同源仅存在无患子科的荔枝、龙眼和红毛丹中,表明该通路可能是在这三个相近物种的共同祖先分离之前进化而来的(图3f)。
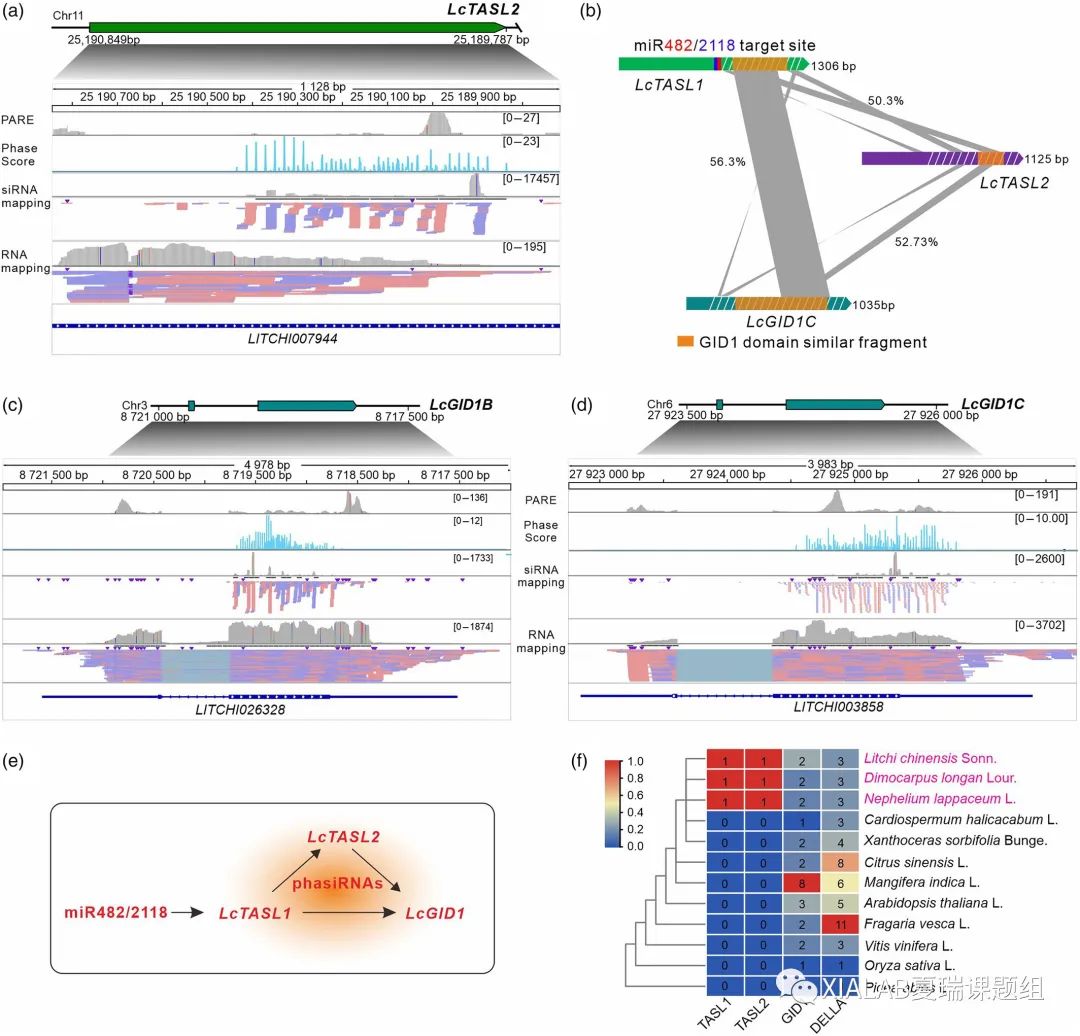
图3 LcTASL1产生的phasiRNA级联触发LcTASL2产生 phasiRNA调控LcGID1
04
miR482/2118-TASL-GID1途径可能参与种子发育
通过表达分析发现LcTASL1、LcTASL2及其产生的phasiRNA在幼果中特异高表达,表明miR482/2118-TASL-GID1途径可能在荔枝果实发育中发挥重要作用(图S10)。进一步采集种子正常发育的大核荔枝品种‘怀枝’(‘HZ’)从未授粉的子房到成熟果实整个发育过程中的果实样本(图4a),通过转录组测序分析发现miR482/2118-TASL-GID1中的LcTASL基因以及LcGID1的表达可能在在种子中可能发挥更重要的作用(图4b)。
随后,本研究对该通路中的sRNA在荔枝种子中的表达进行分析,发现大多数miR482/2118s没有显示出一致的表达模式,这与miR482/2118的不同功能一致(图4c)。而产生于LcTASL/LcGID1的phasiRNA在授粉后显著上调,并在24 DAP达到峰值随后缓慢变化(图4d)。荔枝种子中phasiRNA的高丰度,特别是LcTASL2衍生的phasiRNA高积累,进一步证实了miR482/2118-TASL-GID1途径在种子发育中的作用。
我们同时也对种子败育的焦核荔枝品种‘糯米糍’(‘NMC’)中分析了这些关键基因和sRNA的表达(图S11)进一步表明该途径可能对荔枝的种子发育至关重要。
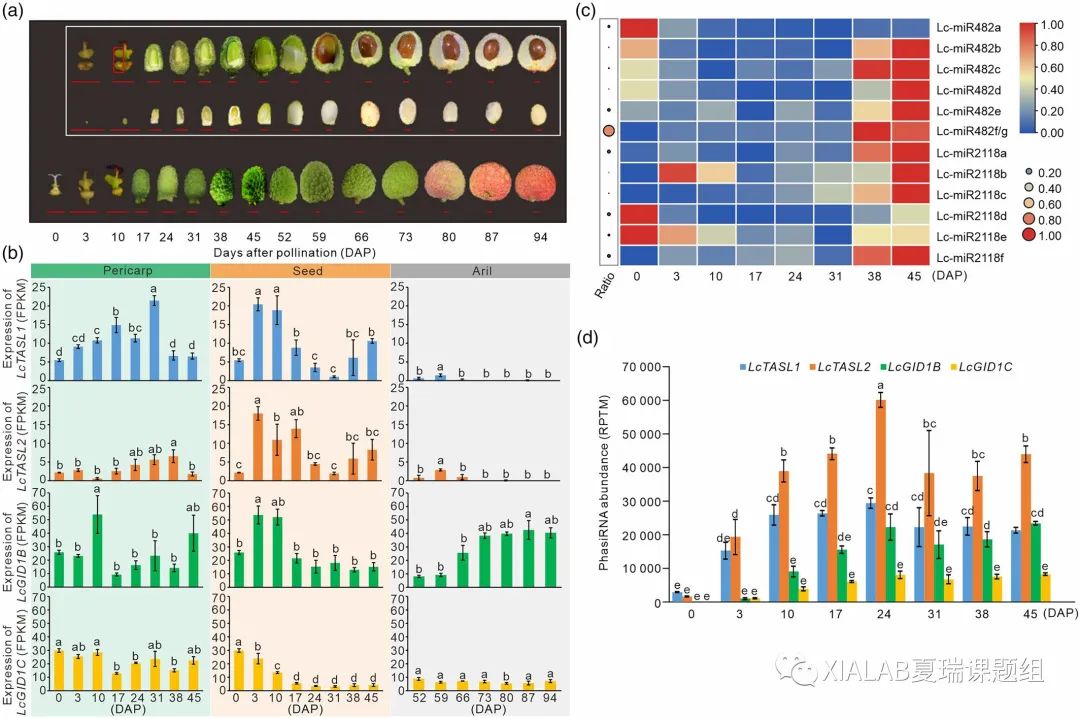
图4 途径在荔枝品种‘HZ’果实发育中关键基因和相关小RNA的表达
05
拟南芥中miR482/2118-TASL-GID1途径影响花果发育
本研究通过在拟南芥中构建Lc-miR482a-LcTASL-AtGID1通路,我们发现OX-LcTASL1和OX-LcTASL2的花丝长度都显著变短(图5a和5b),而OX-LcMIR482a以及LcTASL1 × LcMIR482a没有显著变化。同时,含有LcTASL1的所有基因重组系的果荚显示出不同程度的异常,包括种子败育和较小的荚大小(图5c-e)。观察到过表达的LcTASL1与角果长度和种子数量/角果长度之比呈负相关(图5d和5e),表明构建的途径影响拟南芥的种子和果实发育。基因重组系中的这些缺陷与拟南芥gid1突变体的表型非常相似,后者具有较短的丝和较短的角果,种子较少。
因此,这些结果证明miR482/2118-TASL-GID1通路可能通过调节赤霉素信号传导影响花朵和种子的形成,并最终影响果实的发育。
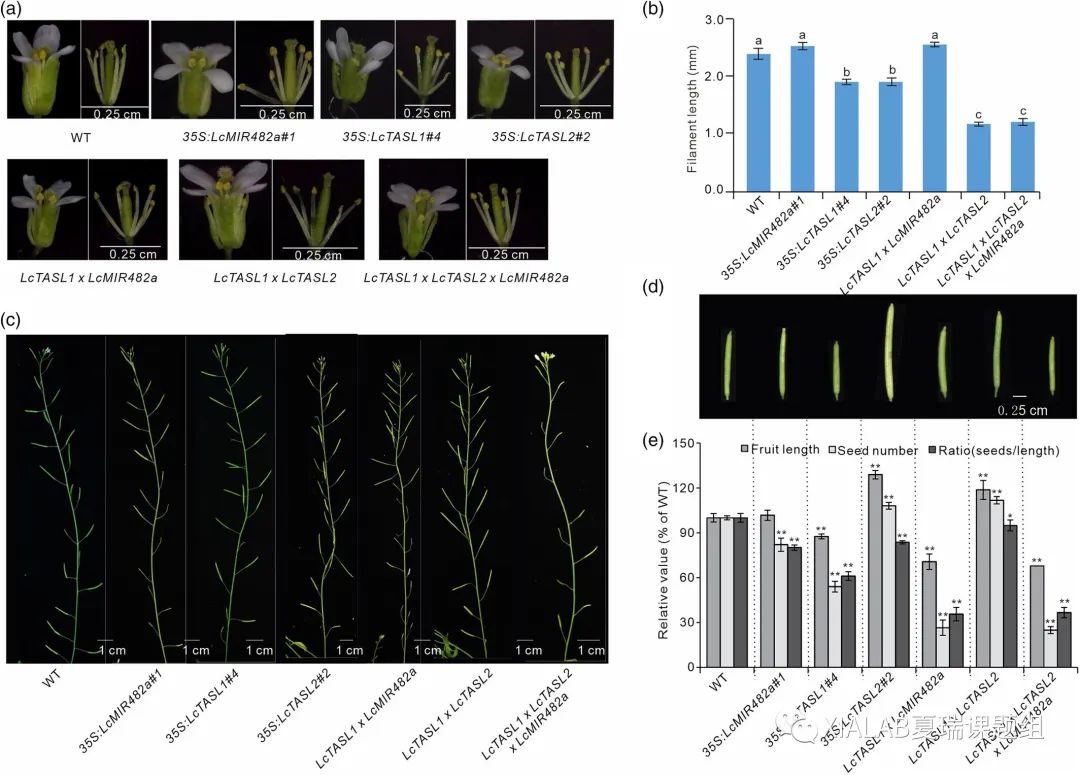
图5 miR482/2118-TASL-GID1途径影响拟南芥开花和果实发育
06
赤霉素响应因子GAMYB反馈调控LcMIR482/2118
通过对LcMIR482/2118s启动子序列分析,发现LcMIR482d和LcMIM482e的启动子含有一类响应赤霉素的R2R3类的MYB基因(GAMYB)结合的GARE基序,而GAMYB是GA传导中的关键基因。对荔枝中鉴定的四个GAMYB基因表达分析发现LcGAMYB33均高于其他三个成员,因此将其作为miR482/2118-TASL-GID途径反馈调控的候选基因(图6a)。对LcGAMYB33进行了DNA亲和纯化测序(DAP-seq)发现其在基因组上有大量的结合位点(图6b),其中结合的核心基序为“TAACTGACT”(e value = 2.0e-212)(图6c)。DAP-seq分析发现LcGAMYB33可以结合在LcMIR482e和LcMIR482f启动子上(图6d和e)。进一步通过电泳迁移阻滞分析(EMSA)和双荧光素酶实验验证了LcGAMYB33通过直接与LcMIR482e和LcMIM482f的启动子结合(图f-h)。
因此,LcMIR482e和LcMIR482f在LcGAMYB33的转录调控下,在荔枝中形成了一个新的反馈调控环。
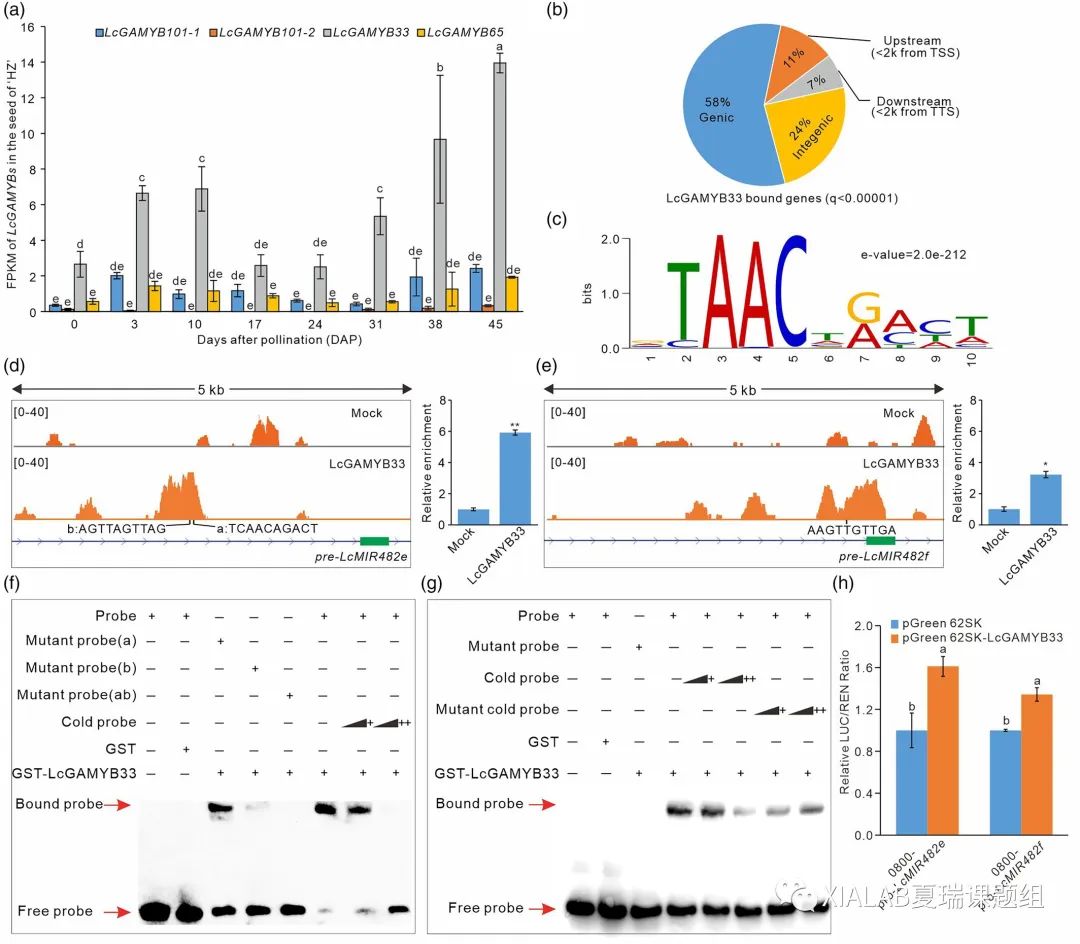
图6 LcGAMYB33调控LcMIR482e 和 LcMIR482f的转录
总的来说,Lc-miR482/2118s通过靶向长链非编码RNA基因LcTASL1,诱导产生大量的21-nt phasiRNA负调控LcGID1s表达从而参与赤霉素信号传导。LcTASL1位点上产生的phasiRNA对LcGID1s的沉默作用可以通过另一个长链非编码RNA基因LcTASL2产生phasiRNA来增强,即miR482/2118-TASL-GID途径(图7)。本研究发现该途径可能与荔枝的种子发育有关。此外,赤霉素信号响应的一个关键因子LcGAMYB33在转录水平上反馈调节MIR482/2118的表达(图7)。这些结果首次证明了miRNA通过触发靶基因产生phasiRNA而直接参与赤霉素信号传导。
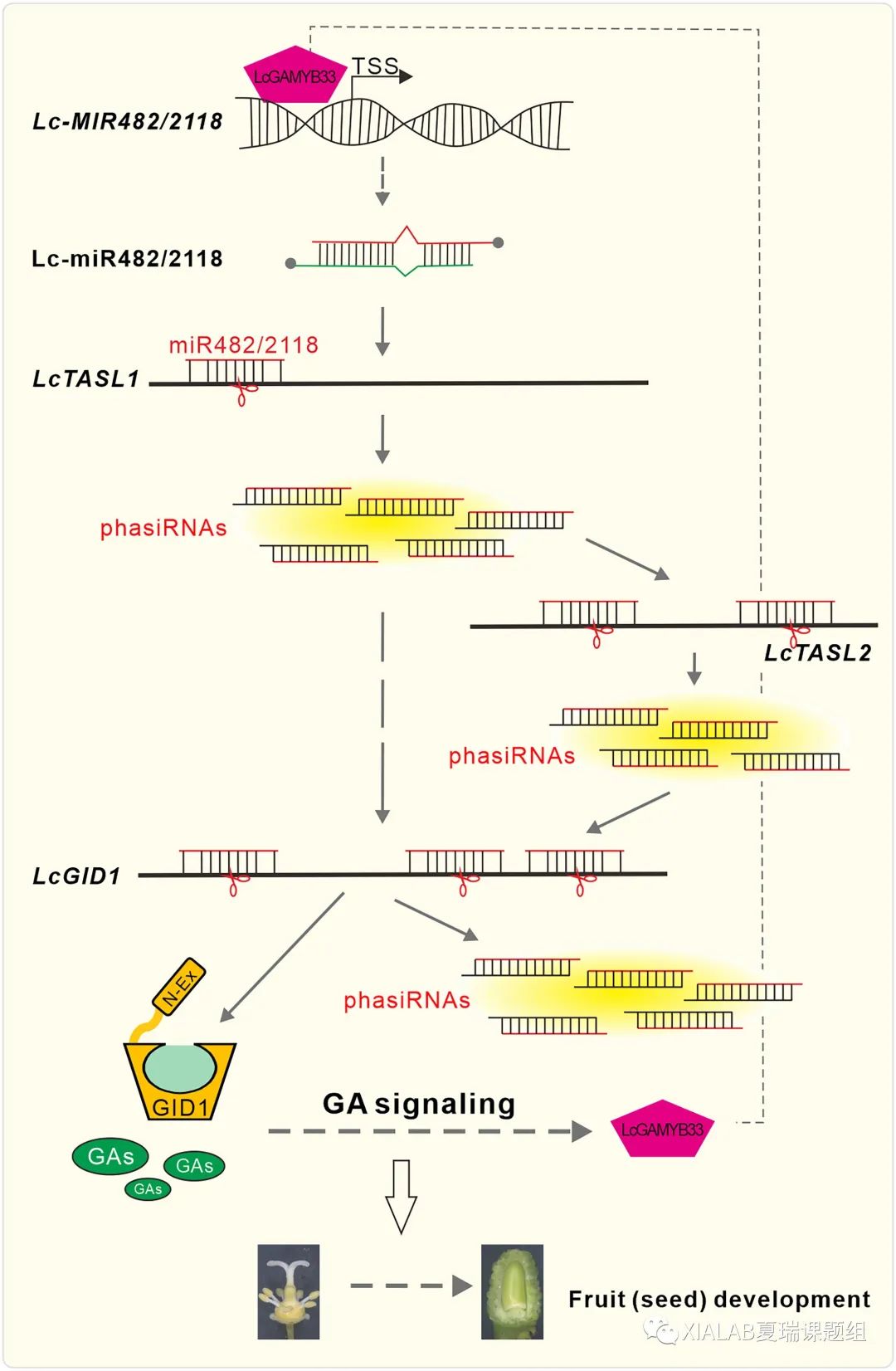
图7 miR482/2118-TASL-GID1 调控通路的模型
本论文以华南农业大学为第一完成单位,华南农业大学园艺学院夏瑞教授、徐婧老师为共同通讯作者,已毕业博士生张艳青为论文第一作者。李建国研究员,副教授赵明磊、郝彦伟、马兴帅、陈程杰、刘元龙,曾灶海实验师,博士研究生胡慧敏和硕士研究生李冠良参与了论文的设计、实验和分析、写作修改。该研究得到了国家自然科学基金(32072547,31872063)、岭南现代农业科学与技术广东省实验室(NZ2021007)以及广东省特支计划(2019TX05N193)的资助。
原文链接:
https://onlinelibrary.wiley.com/doi/10.1111/pbi.14226