
[New Phyologist]华南农大夏瑞课题组综述miR482/2118家族起源,进化及功能多样化
时间:2021-11-18 阅读量:1270
2021年11月11日,华南农业大学夏瑞课题组线上发表了题为“MicroRNA482/2118, a miRNA superfamily essential for both disease resistance and plant development”的综述文章。

miR482/2118是植物中一个古老且相对保守的22nt的miRNA超家族,在植物的生长发育和抗病抗逆过程中发挥重要的作用。该家族成员可以通过触发靶基因产生大量的phasiRNA,而这些phasiRNA即可以顺式作用于其前体基因加强miR482/2118的沉默效应,也可以通过反式作用靶向其它基因,从而扩大miR482/2118沉默作用的效应。
在这篇综述里,作者回顾了miR482/2118自发现以来在起源进化、多样化过程、主要生物学功能等方面的相关研究进展,并对该家族未知的一些功能作用和调控方式提出了相关的展望。综述内容简述如下:
1. 认识miR482/2118超级大家族
miR482/2118是一个22nt的miRNA超家族,其产生是经过经典的miRNA合成通路(Fig. 1a)。作为22nt的miRNA,miR482/2118可以通过“one-hit”的模式触发phasiRNA的产生,从而增加和扩张miR482/2118沉默效应和作用范围。
miR482/2118在植物中有两个主要的类型,miR482型和miR2118型。2005年第一个miR482在杨树中鉴定得到,2008年和2009年陆续在玉米和水稻中鉴定到在生殖器官中特异表达的miR2118。2012年因为两个miRNA家族的序列存在高度相似性,研究人员将miR482和miR2118合并为一个家族,并合称为miR482/2118超家族。
从茎环序列比对可以看出,miR482型通常是“UC”开头,而miR2118型相比于miR482型通常有2nt的移位(Fig. 1b)。一些研究中发现,miR482型和miR2118型的成熟链可能会从同一个前体基因上产生。miR482/2118家族成员的茎环结构第十位都有一个保守的不对称的泡状结构。
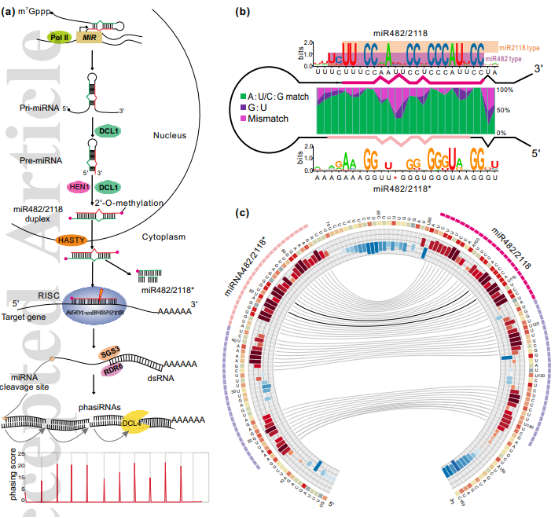
图1. 植物中miR482/2118超家族的生物合成和保守性
根据前体比对的CIRCOS图中可以看出在产生miR482/2118成熟链和星链相邻的下端区域序列高度相似且匹配良好(Fig. 1c),这一段的保守可能是为了保证DCL1精准识别切割,说明该家族很可能是通过从茎到环(base to loop)的剪切方式产生成熟的miRNA。
同时,miR482/2118家族和动物一样,也能在一个位点成簇地存在,比如水稻中18个miR2118的成员聚集在4号和11号染色体上。这些miR482/2118簇可能被转录成单顺反转录本,然后加工成不同的miRNA,表明这些miR482/2118成员可能在在转录水平上受到共同调控。
2. miR482/2118超级大家族的起源和进化
除了2nt的位移,miR482/2118早期的时候被命名为两个不同基因家族的另一原因是它们靶基因的不同。在双子叶植物中miR482/2118主要靶向抗病基因NBS-LRR,而在单子叶植物中则主要靶向长链非编码基因。两者都可以在靶基因上触发大量的phasiRNA的产生,但是前者主要在抗病方面发挥功能,而后者主要是与生殖发育有关。有趣的是,在裸子植物挪威云杉中这两种情况都存在。因此推断,miR482/2118在裸子植物中存在双功能,在双子叶和单子叶植物中进行了功能分化(Fig. 2a)。
miR482/2118最早出现在裸子植物中,对应其靶基因NBS-LRR的极大扩张,表明两者之间可能存在共进化的关系(Fig. 2b)。miR482/2118起源于抗病基因NBS-LRR的反向复制(Fig. 2c),NBS-LRR基因在基因组自发的反向复制中形成完美的茎环结构,在进化过程中不断积累突变,破坏前体的互补性,促成DCL1的识别和加工,最终产生miR482/2118的位点。
一个新的miRNA位点一旦产生,各种选择压力会促进其扩张和功能进化,比如miRNA的靶基因可以驱动miRNA的进化。miR482/2118靶向NBS-LRR基因中功能十分重要的保守结构域P-loop,此结构域在核酸密码子第三位上的序列摇摆性,驱动了miR482/2118家族成员对应位置序列上的多样性(Fig. 2d)。
与单子叶植物相比,这种多样化在裸子植物和双子叶植物中更加明显,也进一步反映了miR482/2118在裸子植物/双子叶植物和单子叶植物之间的功能差异。miR482/2118的多样化与其靶基因NBS-LRR保持同步,这可能是植物平衡防御成本和收益的一种策略。
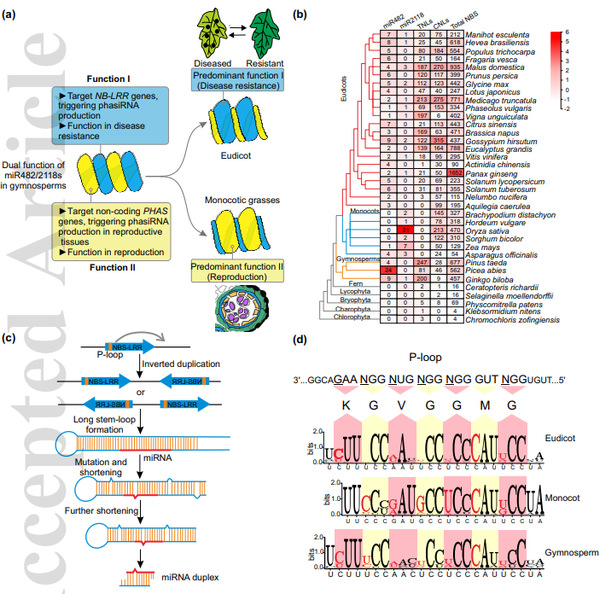
图2. miR482/2118超家族的进化和功能多样化
3. miR482/2118超家族的功能多样化
miR482/2118在裸子植物中的两类主要靶基因在单子叶植物和双子叶植物中出现分化。在双子叶植物中,miR482/2118靶向一类R基因——NBS-LRR基因,并且偏好性地靶向CC类型的NBS-LRR基因。通过靶向NBS-LRR保守结构域P-loop并进行切割(Fig. 3a),miR482/2118家族可以靶向数量庞大的抗病基因家族,这可能是植物中非常实惠和高效的基因调控策略。
同时,miR482/2118切割靶基因NBS-LRR并产生大量的phasiRNA,可以进一步增强miR482/2118对NBS-LRR的沉默效应。这表明双子叶植物中miR482/2118靶向NBS-LRR以及随后产生的phasiRNA参与到植物免疫zig-zag模型中特异的ETI部分(Fig. 3b)。
在单子叶植物中,miR482/2118的主要功能是通过靶向长链非编码基因触发大量的phasiRNA产生,从而参与到生殖过程中(Fig. 3c)。在水稻中,两系杂交中光温敏雄性不育(P/TMS)至关重要。前人研究中发现两个调控光温敏不育的关键的长链非编码基因PMS1T/PMS3都可以被miR2118靶向,并产生大量的phasiRNA,这一通路主要在水稻的花药里发生。两个位点上性状的变化都是由于第二个循环phasiRNA(从miR2118切割位点起)中的1个SNP导致的,说明phasiRNA可能通过某种未知的机制对水稻的育性产生了影响。
目前对于单子叶植物生殖发育过程中的大量phasiRNA的作用机制还不是很清楚。有报道说明,水稻中miR2118靶向长链非编码基因产生的phasiRNA可能是通过不同的机制去发挥功能,如在水稻精细胞和花药壁细胞中不同类型的phasiRNA分别加载到不同的AGO蛋白中发挥作用(Fig. 3c)。
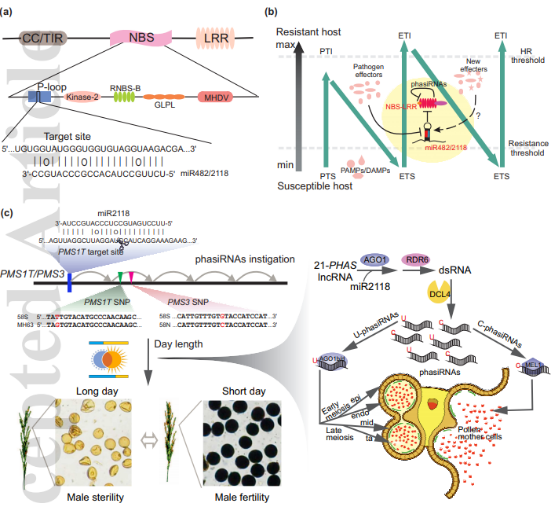
图3. miR482/2118在抗病和生殖生长方面的作用
除了上述的主要功能之外,miR482/2118在物种进化过程中不断获得新的靶基因,从而赋予其在非生物胁迫和其它发育过程中的调控作用。如,在荔枝中miR482/2118靶向能量代谢相关的基因,比如PGK和 Ca2+-ATPase。草莓中新靶基因如F-box可能与果实发育有关。已经有研究表明,在植物的干旱,温度等环境胁迫中有不同的响应。在杨树中发现,miR482/2118靶向4个编码UDP-糖基转移酶的基因,表明其可能在逆境适应中发挥作用。
夏瑞课题组张艳青博士和Muhammad Waseem博士后为该综述的共同第一作者,夏瑞老师通讯作者。南方科技大学翟继先教授也参与了综述的写作,文章得到国家自然科学基金和广东省特支计划等项目的支持,同时感谢美国唐纳德·丹佛斯植物科学中心的 Blake C. Meyers教授在文章写作中提供的宝贵建议。
文章链接:https://nph.onlinelibrary.wiley.com/doi/abs/10.1111/nph.17853